Human Adiponectin DTSet enzyme-linked immunoassay kit
| Specification | 96*5 Test;96T*15 Test |
|---|---|
| Standard Curve Range | 62.50pg/ml-4000pg/ml |
| Standard Curve Gradient | 7 Points/3 Folds |
| Number of Incubations | 2 |
| Detectable sample | Liquid phase sample of soluble substances. For example: serum, plasma, cell culture supernatant, tissue grinding liquid, etc. |
| Sample Volume | 50 μl |
| Type | Not Ready-to-Use |
| Test Duration | 120min |
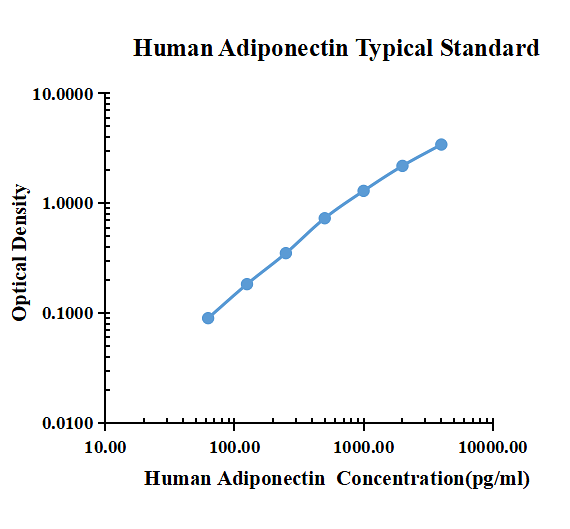
| pg/ml | O.D. | Average | Corrected | |
|---|---|---|---|---|
| 0.00 | 0.0163 | 0.0168 | 0.0166 | |
| 62.50 | 0.1094 | 0.1034 | 0.1064 | 0.0899 |
| 125.00 | 0.1902 | 0.2094 | 0.1998 | 0.1833 |
| 250.00 | 0.3602 | 0.3745 | 0.3674 | 0.3508 |
| 500.00 | 0.7332 | 0.7577 | 0.7455 | 0.7289 |
| 1000.00 | 1.2980 | 1.3210 | 1.3095 | 1.2930 |
| 2000.00 | 2.1300 | 2.2660 | 2.1980 | 2.1815 |
| 4000.00 | 3.2680 | 3.5890 | 3.4285 | 3.4120 |
Product Features
- Optimized capture and detection antibody pairings with recommended concentrations save lengthy development time
- Development protocols are provided to guide further assay optimization
- Assay can be customized to your specific needs
- Economical alternative to complete kits
Kit Content
- Capture Antibody
- Detection Antibody
- Recombinant Standard
- Streptavidin conjugated to horseradish-peroxidase (Streptavidin-HRP)
Other Reagents Required
DTSet Ancillary Reagent Kit (5 plates): containing 96 well microplates, plate sealers, substrate solution, stop solution, plate coating buffer (PBS), wash buffer, and assay buffer.
- 96 well microplates: YOUKE Life, Catalog # DSEP01. Plate Sealers: YOUKE Life, Catalog # DSSF01.
- Coating Buffer: 137 mM NaCl, 2.7 mM KCl, 8.1 mM Na2HPO4, 1.5 mM KH2PO4, pH 7.2-7.4, 0.2μm filtered . YOUKE Life, Catalog # DSCB01.
- Blocking Buffer: YOUKE Life, Catalog # DSBB01.
- Wash Buffer: 0.05% Tween® 20 in PBS, pH 7.2-7.4. YOUKE Life, Catalog # DSWB01.
- Assay Buffer: 0.5% BSA,0.05% Tween® 20,PBS Solution.YOUKE Life, Catalog # DSAB01
- Substrate Solution: Tetramethylbenzidine. YOUKE Life, Catalog # DSTS01.
- Stop Solution: 0.5mol/ml H2SO4. YOUKE Life, Catalog # DSSS01.
Product Data Sheet
Background: Adiponectin
Adiponectin, alternately named Adipocyte complement-related protein of 30 kDa (Acrp30), adipoQ, adipose most abundant gene transcript 1 (apM1), and gelatin-binding protein of 28 kDa (GBP28), is an adipocyte-specific, secreted protein with potential roles in glucose and lipid homeostasis. Circulating Adiponectin levels are high, accounting for approximately 0.01% of total plasma protein. Adiponectin contains a modular structure that includes an N-terminal collagen-like domain followed by a C-terminal globular domain with significant sequence and structural resemblance to the complement factor C1q. Although they share little sequence identity, similar threedimensional structure and certain conserved amino acid residues suggest an evolutionary link between the C1q-like domain of Adiponectin and members of the TNF superfamily. Adiponectin assembles into different complexes including trimers (low molecular weight), hexamers (middle molecular weight), and higher order oligomeric structures (high molecular weight) that may affect biological activity. Adiponectin is induced during adipocyte differentiation and its secretion is stimulated by insulin. Two receptors for Adiponectin, termed AdipoR1 and AdipoR2, have been cloned. Although functionally distinct from G-protein-coupled receptors, the genes encode predicted proteins containing 7 transmembrane domains. AdipoR1 is highly expressed in skeletal muscle, while AdipoR2 is primarily found in hepatic tissues.
Injection of Adiponectin into non-obese diabetic mice leads to an insulin-independent decrease in glucose levels. This is likely due to insulin-sensitizing effects involving Adiponectin regulation of triglyceride metabolism. A truncated form of Adiponectin (gAdiponectin) containing only the C-terminal globular domain has been identified in the blood, and recombinant gAdiponectin has been shown to regulate weight reduction as well as free fatty acid oxidation in mouse muscle and liver. The full-length recombinant Adiponectin protein is apparently less potent at mediating these effects. The mechanism underlying the role of Adiponectin in lipid oxidation may involve the regulation of expression or activity of proteins associated with triglyceride metabolism including CD36, acyl CoA oxidase, AMPK, and PPAR gamma.
Although Adiponectin-regulation of glucose and lipid metabolism in humans is less clear, similar mechanisms may also be in place (15. A negative correlation between obesity and circulating Adiponectin has been well established, and Adiponectin levels increase concomitantly with weight loss. Decreased Adiponectin levels are associated with insulin resistance and hyperinsulinemia, and patients with type-2 diabetes are reported to exhibit decreased circulating Adiponectin. Thiazolidinediones, a class of insulin-sensitizing, anti-diabetic drugs, elevate Adiponectin in insulin-resistant patients. In addition, high Adiponectin levels are associated with a reduced risk of type-2 diabetes. Using magnetic resonance spectroscopy it has been demonstrated that intracellular lipid content in human muscle negatively correlates with Adiponectin levels, potentially due to Adiponectin-induced fatty acid oxidation.
Adiponectin may also play anti-atherogenic and anti-inflammatory roles. Adiponectin plasma levels are decreased in patients with coronary artery disease. Furthermore, neointimal thickening of damaged arteries is exacerbated in Adiponectin-deficient mice and is inhibited by exogenous Adiponectin. Adiponectin inhibits endothelial cell expression of adhesion molecules in vitro, suppressing the attachment of monocytes. In addition, Adiponectin negatively regulates myelomonocytic progenitor cell growth and TNF-alpha production in macrophages.
